![[Vaccine] 世界で開発された 新型コロナウイルス用ワクチンの有効性 – Pfizer, Moderna, Astrazeneca, Sinovac [2021/06/15]](https://harikiri.diskstation.me/wp-content/uploads/2020/04/CEEBD8EE-9D2E-4F43-B074-64ABE84F4C6B.jpeg)
はじめに
変異株は、イギリス株に加えインド株(デルタ株)が、感染力が良いため、感染拡大の問題が明らかになってきました。イギリスでは、国民全体にワクチン接種を早期に完了させることを戦略として、2回摂取までの間隔を長くすることで感染の進行を抑えてきました。ここに来て、インド由来の変異株であるデルタ株の感染が増加しており、市中感染においてデルタ株に殆どが置きかわっているとされます。2回摂取の必要性が明らかに成りつつあります (NHK News, 2021/06/14)。
- 2021/05/23, インド変異株(デルタ株)に対する有効性について、1摂取では、AstrazenecaとPfizerとも33%, 2回摂取でAstrazenecaは60%, Pfizerは88%であると、イングランド公衆衛生庁(PHE)が発表, BBC News(ファイザー製とアストラゼネカ製ワクチン、2回接種でインド型変異株に有効=英研究, 2021/05/23))
各社のワクチン
新型コロナウイルスに対するワクチンは、世界で開発が進められています。最も進んでいるワクチンは、第3相の臨床試験の結果が既に得られており、その有効性が確認されています。
開発を先行している製薬会社として、アメリカのPfizerとModerna、およびイギリスのAstrazenecaの3社の有効性について比較した表を以下に示します (ひるおび, TV, 2020/12/01, 一部改変)。
表1. mRNAワクチンとウイルス・ベクター・ワクチンの比較
| 項目 | Pfizer | Moderna | Astrazeneca | Jannsen |
| 企業国 | アメリカ | アメリカ | イギリス | |
| ワクチンタイプ | mRNA | mRNA | Adenovirus Vector | mRNA |
| 有効性 (第3相臨床試験) | 95% | 94.1% | 平均70% 最高90% | 重症化への抑制率は、80%程度 |
| 摂取 | 2回 | 2回 | 2回 | 1回 |
| 保存条件 | -80 ~ -60℃, ~ 半年 2 ~ 8℃ (5days) | – 20℃, ~ 半年 2 ~ 8℃ (30days) | 2 ~ 8℃ (半年以上) | |
| 日本への供給 | 国内での審査/承認は、早くても2021/02~03, ~ 2021/06 (6,000万人分) | ~ 2021/09 (2,500万人分) | 2021 ~ (6,000万人分) | N/A |
| アメリカでの提供 | 2020/12/11, FDAは緊急使用を認めた(emergency use authorization (EUA)*1) | 2020/12/01, アメリカでの緊急使用を申請 | ||
| 提供国 | イギリス(2020/12/07= (予想)), EU | U.S. | EU | スペイン |
| 臨床 | 2020/11/8 | |||
| 副作用 | 血栓が危惧されている |
*1: EUAのマイルストーンを達成
- 一次有効性分析は、BNT162b2が初回投与の28日後にCOVID-19に対して95%有効であることを示した; 170の確認されたCOVID-19の症例が評価され、プラセボ群で162例、ワクチン群で8例が観察された。
- 有効性は、年齢、性別、人種、民族の人口統計全体で一貫していた
- 65歳以上の成人で観察された有効性は94%以上
- 米国食品医薬品局(FDA)が緊急使用許可(EUA)に要求する安全性データのマイルストーンが達成された
- データは、43,000人以上の参加者が登録されたすべての集団でワクチンが十分に許容されたことを示している。
- 重大な安全上の懸念は観察されなかった。
- 頻度が2%を超える唯一のグレード3の有害事象は、3.8%の倦怠感と2.0%の頭痛であった
- pfizerは、EUAのために数日以内にFDAに提出し、世界中の他の規制当局とデータを共有することを計画している
- 両社は、2020年に世界で最大5,000万回のワクチンを製造し、2021年末までに最大13億回のワクチンを製造する予定
- pfizerは、ワクチンを世界中に配布するための豊富な経験、専門知識、および既存のコールドチェーンインフラストラクチャに自信を持ってる
PFIZER AND BIONTECH CONCLUDE PHASE 3 STUDY OF COVID-19 VACCINE CANDIDATE, MEETING ALL PRIMARY EFFICACY ENDPOINTS
https://www.pfizer.com/news/press-release/press-release-detail/pfizer-and-biontech-conclude-phase-3-study-covid-19-vaccine
1回の接種で済むジョンソン・エンド・ジョンソンの新型コロナワクチンは米国で3月に配布開始の見込み
https://jp.techcrunch.com/2021/01/30/2021-01-29-johnson-johnsons-covid-19-vaccine-is-85-effective-against-severe-cases-and-66-effective-overall-per-trial-data/?guccounter=1&guce_referrer=aHR0cHM6Ly93d3cuZ29vZ2xlLmNvLmpwLw&guce_referrer_sig=AQAAAHWrulzT5dtNkepyOEfFTC-ncFJkYJ_Qu2Dr4Hsfximdft7bNNibxg4ZE-mkWPLZUrbi2CHc68WnFCOudaD31tU8K7fPJJvbVHlmP7KyKY114TuFC4ZnirWqr58xYKq2jlg2XE_ve15FNnkOkdEJRKuzXCJ2YK5kMCfejJNGtfdT
評価
当初、僕が最初に調査した時は、核酸ワクチンと比較して、ウイルス・ベクターの方が免疫賦活化効果が高いのではないかという印象がありました。ところが、今回の2種類のmRNAワクチンと1種類のウイルス・ベクター・ワクチンとで、有効性に違いが見られました。mRNAワクチンの方が高いという結果です(表1)。いずれのモダリティーも新しいモダリティーであり、世の中には、まだ存在しない医薬品です。今後も、それぞれのモダリティーにおいても改良が加えられた製品が出て来ますが、ワクチン効果としての有効性が、本当にmRNAワクチンに軍配が上がるのかは、もう少し待ってみようと思います。
*有効性: ワクチンを摂取しても感染するヒトがいますし、当然、ワクチンを打っていなければ、その感染率は更に高いはずです。有効性とは、ワクチンを摂取した群での感染者数が5人であったとき、ワクチンを摂取していない群での感染者数が100である場合に、有効性は、95%になります。100人、5人では、統計学的に信頼性が低いので、データとして信頼性が高くなるように、第3相臨床試験では、全体の試験に参加するボランティアは4万人程度、そこから、偽薬とワクチンのそれぞれの群に2万人ずつに群分けして、臨床試験が実施されます。それぞれの群で、発症した患者数を比較します。
その他
CoronaVac (by SinoVac)
- 中国、シノバックのCOVID-19ワクチン(CoronaVac)は、Vero細胞で増やして不活化処理した不活化ワクチンです。ブラジルのサンパウロで使用され、その有効性は、50.4%であると報じられています。
- Virus strain (ウイルス株) : CZ02
- Adjuvant (アジュバント) : Aluminum Hydroxide
- Excipients (添加剤) : リン酸水素2Na (12水和物), リン酸2水素Na, NaCl
- milky-white suspension
NIKKEI Asia – Sinovac vaccine disappoints at 50.4% efficacy: latest Brazil data – 2021/01/13
https://asia.nikkei.com/Spotlight/Coronavirus/Sinovac-vaccine-disappoints-at-50.4-efficacy-latest-Brazil-data
COVID-19 Vaccine (Vero Cell), Inactivated (Brief Edition), 薬剤名 : CoronaVac
https://www.fhb.gov.hk/download/our_work/health/201200/e_PI_CoronaVac_brief.pdf
tozinameran (by Pfizer-BioNTech)
- PfizerのCOVID-19ワクチンの副作用のうち、アナフィラキシーショックは、1/100万人と通常のワクチンに比べて若干高いが、死亡例は報告されていない。日本でも投与が開始間近かとなり、投与後に通常15分、アナフィラキシー既往者の場合は30分の観察の実施により万全が尽くされる(2021/02/15)
- Brand name : Pfizer-BIoNTech COVID-19 Vaccine
- Company name : BioNTech Manufacturing GmbH
- Ingredient : tozinameran
Pfizer-BioNTech COVID-19 Vaccine (tozinameran) – Government of Canada –
https://covid-vaccine.canada.ca/pfizer-biontech-covid-19-vaccine/product-details
編集履歴
2020/12/01 Harikiri(Mr) 2020/12/02 追記(評価、有効性) 2021/01/13 シノバック 2021/05/11 追記 (SinoVacとPfizer-BioNTechのCovid-19ワクチンの薬剤情報) 2021/06/15,追記 (イギリスでは、インドの変異株は置き換わっている)
![[Biologics] 医薬品製造における工程別の名称 : 原料からAPI(有効成分) / DS (原薬),バイオ医薬と低分子医薬での比較 [2025/04/10改定]](https://harikiri.diskstation.me/wp-content/uploads/2021/02/80F3D755-3019-49C6-BDAE-47D137C87826.jpeg)
![今日の英語 – 質問があればご連絡ください – please let me know … [2020/11/24]](https://harikiri.diskstation.me/wp-content/uploads/2020/11/6E416B35-8F79-4004-B7B3-952A457EA637.jpeg)
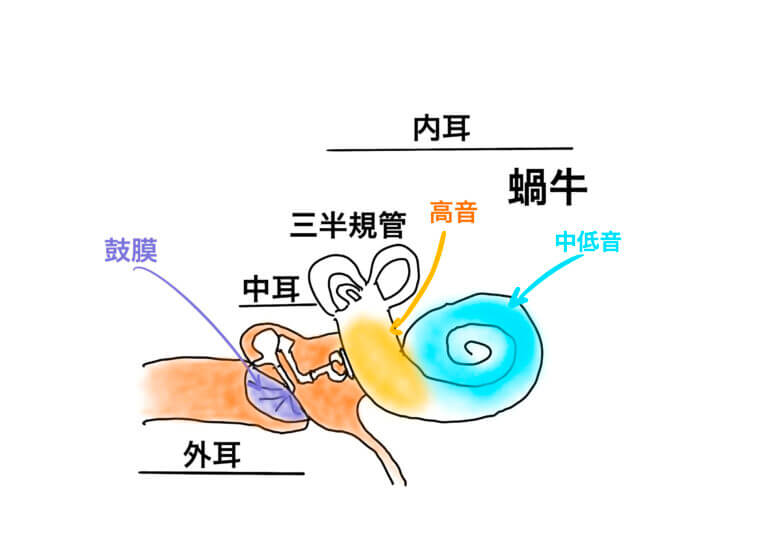
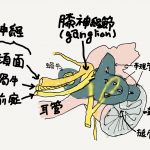
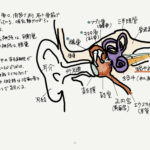
![[健康-偉人] 難聴から「ベートーべン; Beethoven」を学ぶ – [2020/11/23]](https://harikiri.diskstation.me/wp-content/uploads/2020/09/0E1A8FDA-ABDA-4D42-811D-BE0041CBB6CB.jpeg)


![[アニメ] 暗殺教室 (2015,2016) – 中学校教師のアナロジー – 忘れかけていた志(こころざし)が、蘇(よみがえ)る感覚 [2020/11/22]](https://harikiri.diskstation.me/wp-content/uploads/2020/11/1E9497BB-C537-4A01-AD66-7FED5256B8FE.jpeg)
![[Cafe] STARBUCKS in 枚方 蔦屋書店 でblogメンテ [2020/11/21]](https://harikiri.diskstation.me/wp-content/uploads/2020/11/C59FDB78-9E82-4E05-826A-19D858A1B4BE.jpeg)


![気になる企業 ベクタービルダー /デザイン/受託/開発/GMP製造 [2020/11/21]](https://harikiri.diskstation.me/wp-content/uploads/2019/10/B4F24978-42E9-477E-92B0-94605AEBEAB4.jpeg)
![今日の英語 – post/after – 後 – ネイティブは結構、postも使う [2025/04/17]](https://harikiri.diskstation.me/wp-content/uploads/2020/11/F12C8F86-A0B0-4334-9DAD-E75022893870.jpeg)
![[WordPress] AMP対応化で、使用しているその他のプラグインによるエラーが出ないようにするために – プラグイン別 [2020/11/16]](https://harikiri.diskstation.me/wp-content/uploads/2020/02/0AADC961-0278-4A67-B770-47E920676DAD.jpeg)