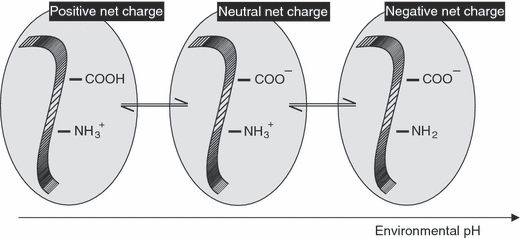
以下,文献によるとウイルスの表面電荷は,およそ酸性から中性域にあります(2.1~8.3).平均値は,5.0±1.3です.ウイルス精製では,陰イオン交換体クロマトグラフィ(AEX)による吸着/溶出で可能です.等電点とは,環境pHに応じて電気的に中性となるpHを等電点(pI or IEP: isoelectric point)と言います.今回紹介する文献では,公開されているウイルスのIEPを再度検証しています.1938年以降に行われた104のウィルスのIEPのデータがまとめられています.
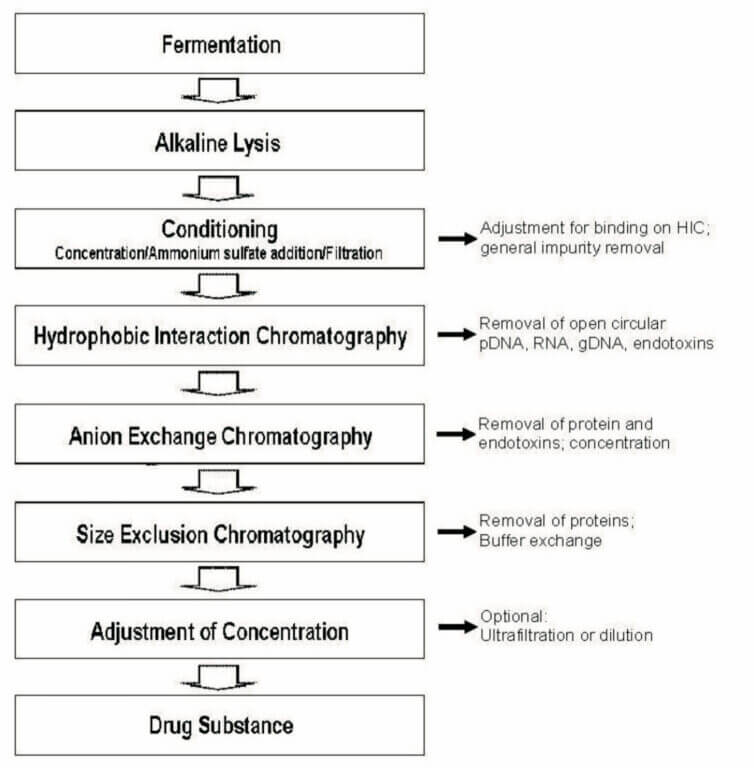
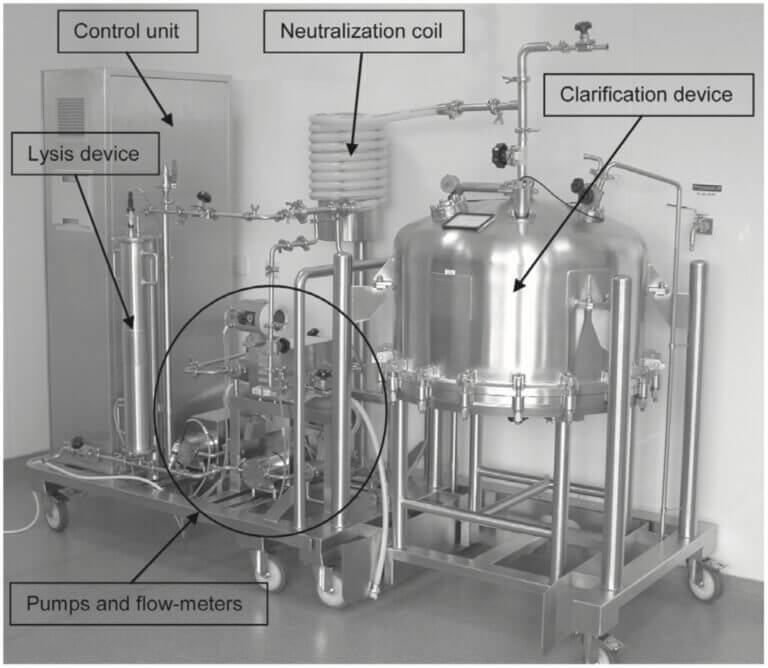
plasmid DNA (pDNA)の製造で最もクリティカルな工程は、その抽出です。pDNAを生産する細胞にE.coliを用いた場合、E.coliのgenomeと目的のpDNAを効率よく分離抽出することが重要です。pDNAは、スーパーコイル(輪ゴムが更によじれている物をイメージするとわかりやすい)となっており、E.coliのgenomeとは物理的強度が異なることを利用して、アルカリ抽出やガラスピーズのミル抽出が一般的に行われます。
5) Hot-Alkaline DNA Extraction Method for Deep-Subseafloor Archaeal Communities (2014)
アルカリ処理と加熱を使用した新しいDNA抽出方法をの検討。 1 M NaOHで98°Cで20分間処理すると、さまざまな深さで収集された海底下の堆積物サンプル中の微生物細胞の98%以上が破壊されました。 しかし、DNAの完全性試験では、このような強アルカリ性および熱処理により、DNA分子がPCRでは増幅できない短い断片に切断されることが示されました。
8) US Patent for Plasmid DNA extraction process Patent – Justia Patents – プラスミドDNA抽出プロセス – FujifilmDiosynth Biotechnolgies UK Limited特許 (2010/07/29出願、US特許8889852)
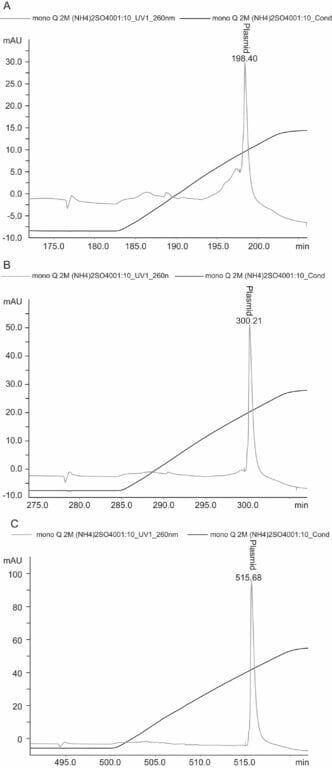
Figure 3. Plasmid purity analysis by HPLC. Plasmid purity was determined by anion-exchange HPLC on a MONO QTM 5/50 GL column. A linear gradient was performed at 0.5 mL/min by increasing the NaCl 0–1 M in 40 mM Tris/HCl, pH 8.0 for 10 CV at a flow rate of 0.5 mL/min. (A) plasmid sample purified by 2 M ammonium sulfate, (B) plasmid sample purified by 2.5 M ammonium sulfate, (C) plasmid sample purified by 3 M ammonium sulfate.
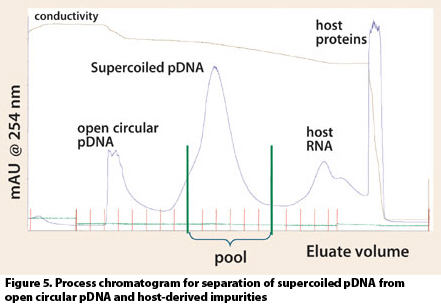
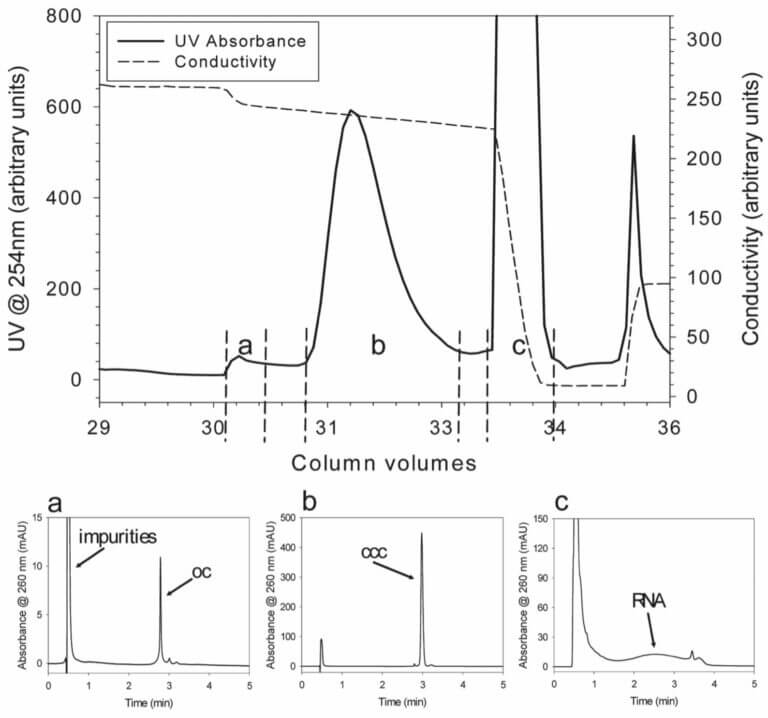
Top: Elution profile of the HIC step (pRZ-hMCP1, 4.9 kbp). Main fractions are indicated by dashed lines. Bottom: Analyti- cal HPLC chromatograms of the HIC fractions; a) the first HIC peak contains impurities and open circular (oc) pDNA; b) the second HIC peak contains mainly supercoiled (ccc) pDNA; c) residual RNA and further im- purities elute when conductivity is suddenly strongly decreased. More strongly bound material is washed out of the column dur- ing regeneration (peak at the end of the HIC chromatogram
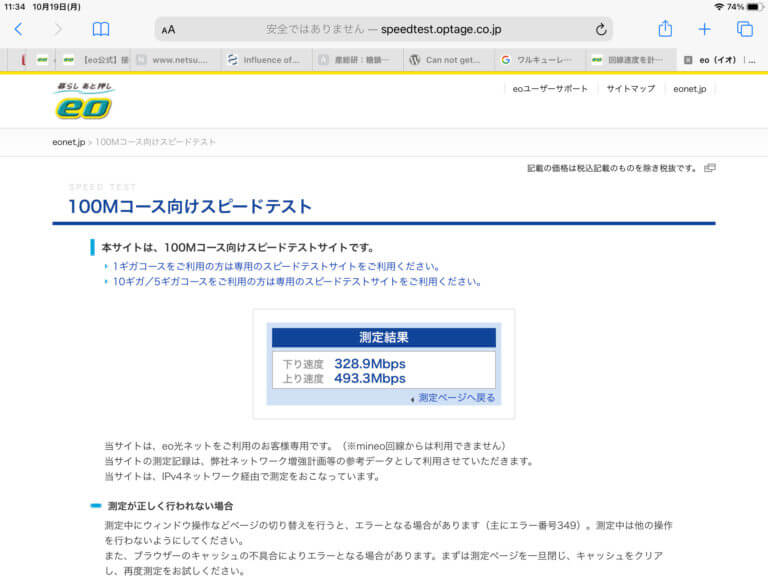
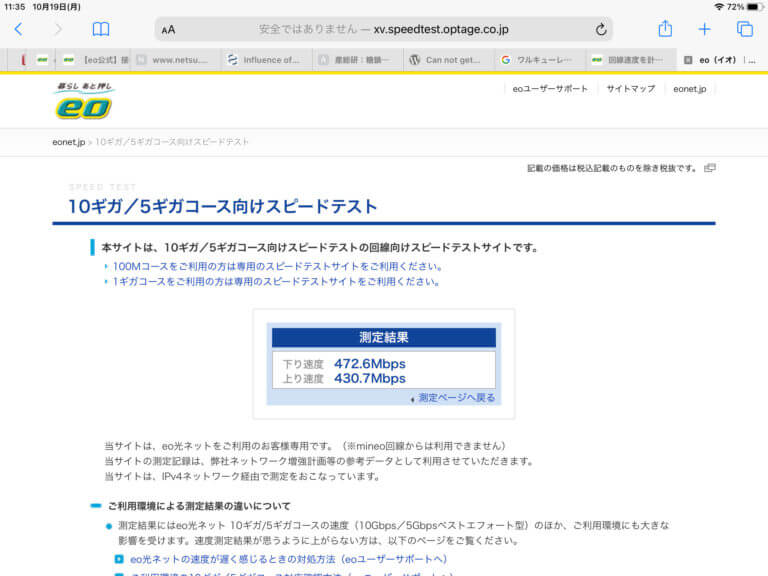
Nintendo Switchの回線テストを実施したとき、まず、インターネットの接続状況を判定し、正常であれは、NATタイプ、ダウンロード速度およびアップロード速度を測定します。それぞの値は、通常、~70kbpsと~30kbpsです。この場合、RT2600acのネットワークモニターでのピーク速度は、ダウンロードで300KB/s、アップロードで6KB/sです。一方、ダウンロードで150KB/sを下回ってくるとと、この測定の段階でエラー(接続不良)を報告してきます。
Nintendo Switchでの回線テストを実施しなくても、インターネットの接続は、断続的に接続/切断を繰り返すことを観察できているので、ゲームによる大容量の通信が原因でインターネットの回線が切断されているとは考えられず、eo光側の問題であると思われます。ただ、RT2600acのハードウェア的な問題である可能性も排除できていません。この問題については、もう一台RT2600acを準備してから検討する予定です。
編集履歴
2020/10/19 Mr.Harikiri
2021/03/27 ホームネットワークのメンテナンスとしてELECOM Laneed 6A型を1m x 2, 2 m x 3, 3m x 1を購入したので、この機に文言整備、CAT5,6および7の違いについて、教えて!goo、blue_plusさんの回答を参照.
2021/03/30 追記 (eo光の「回線テスト」について)
当サイトでは、利便性向上や閲覧追跡にGoogle・他提携サービスによりCookieを使用しています。サイトの閲覧を続けた場合Cookieの使用に同意したことになります。.理解しました拒否Read More
プライバシーポリシー
Privacy Overview
This website uses cookies to improve your experience while you navigate through the website. Out of these, the cookies that are categorized as necessary are stored on your browser as they are essential for the working of basic functionalities of the website. We also use third-party cookies that help us analyze and understand how you use this website. These cookies will be stored in your browser only with your consent. You also have the option to opt-out of these cookies. But opting out of some of these cookies may affect your browsing experience.
Necessary cookies are absolutely essential for the website to function properly. This category only includes cookies that ensures basic functionalities and security features of the website. These cookies do not store any personal information.
Any cookies that may not be particularly necessary for the website to function and is used specifically to collect user personal data via analytics, ads, other embedded contents are termed as non-necessary cookies. It is mandatory to procure user consent prior to running these cookies on your website.
![[用語] パーキンソン病](https://harikiri.diskstation.me/myblog/wp-content/uploads/2021/02/80F3D755-3019-49C6-BDAE-47D137C87826.jpeg)
![[Bio-Edu] ウイルスの等電点(pI) – 組換えAAVでは,empty/fullの違いでpIは異なる – 分離精製は工業化の課題である[2021/02/23]](https://harikiri.diskstation.me/myblog/wp-content/uploads/2020/11/9AF42F79-4BAD-470D-B857-78DB7B3E9802.jpeg)
![[アニメ] 氷菓 (2012) – 高校生の文化祭はこう楽しむべきだったのか – 今更に擬似体験して満足する [2020/11/04]](https://harikiri.diskstation.me/myblog/wp-content/uploads/2020/11/896E1178-29A9-48DC-8A23-55DBEEBEF3E5.jpeg)
![[アニメ] ザ・サード〜蒼い目の少女〜 (2006) – 3つ目の眼を額にもつ「ザ・サード」が人類を管理している世界 [2020/10/20]](https://harikiri.diskstation.me/myblog/wp-content/uploads/2020/10/1FB95525-9A17-482A-8715-9C070F226F58.jpeg)
![[アニメ] 終わりのセラフ – 人対吸血鬼 – 大人の事情と子供の理想は相ゆずれない [2020/10/28]](https://harikiri.diskstation.me/myblog/wp-content/uploads/2020/10/CB823037-BF82-4356-8881-89D60DAFA8E4.jpeg)
![今日の英語 – we are excited to announce ~ – 報告できることを嬉しく思っている表現 [2020/10/28]](https://harikiri.diskstation.me/myblog/wp-content/uploads/2020/11/F12C8F86-A0B0-4334-9DAD-E75022893870.jpeg)
![[アニメ] ヒロイック・エイジ – 宇宙戦闘も迫力、エイジの純粋な存在が物語を作っていく [2020/10/20]](https://harikiri.diskstation.me/myblog/wp-content/uploads/2020/10/06F6F38C-502D-4345-892F-A97C75224C0B.jpeg)
![[Bio-Edu] Plasmid DNA (pDNA)のデザイン及び、その製造方法に関する調査 [2020/12/24]](https://harikiri.diskstation.me/myblog/wp-content/uploads/2020/10/15419700-C78D-4749-B076-A287382D4681.jpeg)
![[用語] HEK293細胞 – その他派生種の概説 [2020/12/22]](https://harikiri.diskstation.me/myblog/wp-content/uploads/2020/07/52A14BA1-7FA5-4C9E-BA19-8AB55D61952D.jpeg)
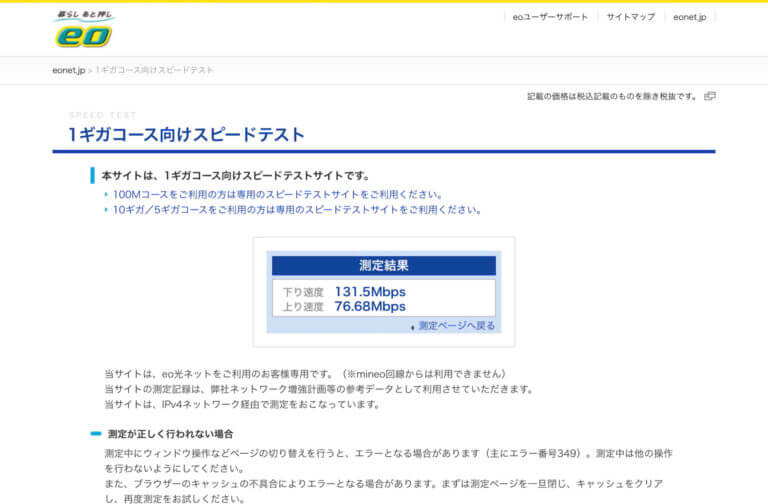
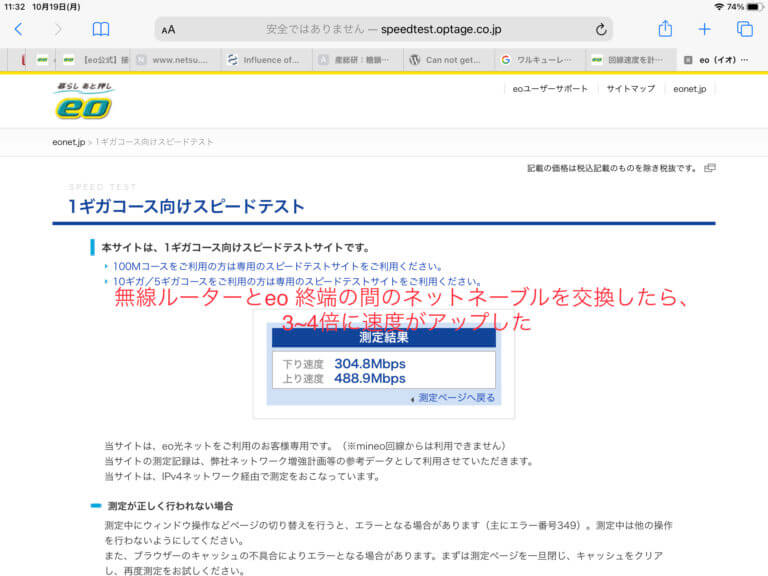
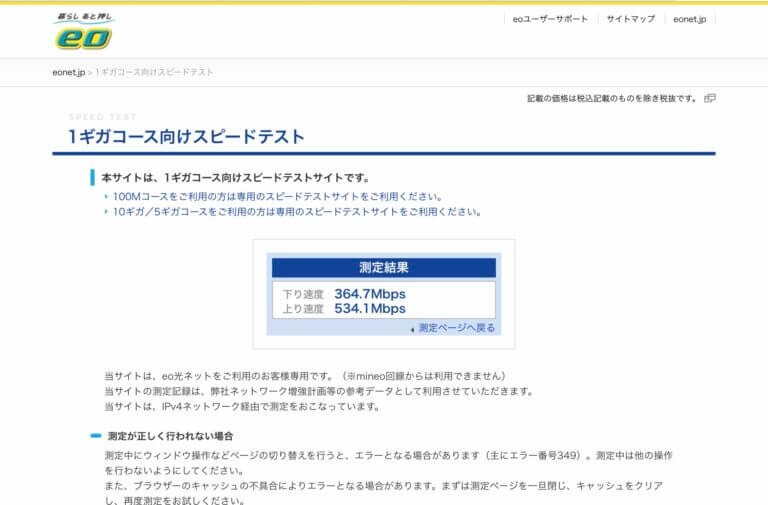
![[Synology] 以前からeo光(1Gコース)なのに速度が遅く感じられていた(100Mbps) – 6A型(CAT6)のネットケーブルで改善 (300 ~ 500Mpbs) [2021/03/30]](https://harikiri.diskstation.me/myblog/wp-content/uploads/2021/02/BBA80493-481A-4463-8AC4-21D8743F7E20.jpeg)